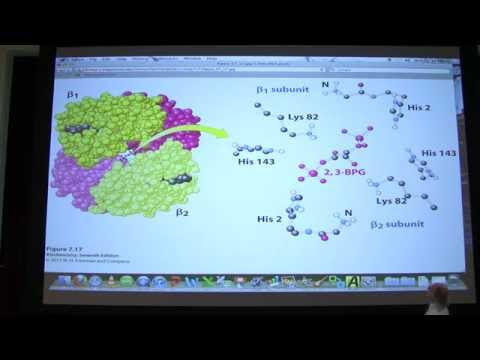
B1 中級美國腔##08生物化學血紅蛋白講座為Kevin Ahern的BB 450/550。 (#08 Biochemistry Hemoglobin Lecture for Kevin Ahern's BB 450/550)00李承育 發佈於 2021 年 01 月 14 日更多分享分享收藏回報影片單字沒有此條件下的單字stateUS /stet/・UK /steɪt/n. (c./u.)州;狀態adj.政府的;州的v.t.陳述,說明,聲明A1 初級多益初級英檢更多 structureUS /ˈstrʌk.tʃɚ/・UK /ˈstrʌk.tʃə/n. (c./u.)結構;建築物v.t.構成;組織A2 初級多益中級英檢更多 proteinUS /ˈprəʊˌtiːn/・UK /ˈprəʊti:n/n. (c./u.)蛋白質B1 中級中級英檢更多 moleculeUS /ˈmɑlɪˌkjul/・UK /ˈmɒlɪkju:l/n.分子B2 中高級高級英檢更多 使用能量解鎖所有單字解鎖發音、解釋及篩選功能解鎖